
Ⅰ. 서 론
벼의 생육기간은 보통 4월에서 10월 사이인데, 이 기간 중에 우리나라 기상조건상 벼의 생육 초, 중기에 긴 장마기간과 중복되어 일조부족현상이 빈번하게 나타나고 있다. 지구의 기상환경의 악화로 인하여 여름철의 장마기간이 길어지어 저온 및 일조 부족으로 인하여 쌀 생산량이 급격히 감소하였고, 여름철의 긴 장마로 인한 일조량의 감소가 나타났으며, 최근 일조부족현상이 출현한 것으로 알려져 있다. 특히, 벼는 재배적 특징상 우리나라의 전역 어느 곳에서도 재배가 가능하여 태백고냉지대(평창, 영월, 정선, 태백 등), 태백준고령지대(양구, 인제, 홍천, 제천 등), 소백산간지대(괴산, 보은, 옥천, 영동), 노령소백산간지대(무주, 진안, 임실, 남원 등), 영남내륙산간지대(영천, 상주, 안동, 진안,금천) 등 중산간지역에서도 널리 재배되고 있으나, 이들 지역 대부분의 논은 계곡이나 산기슭에 위치하고 있는 관계로 산 능선에 의하여 오전 또는 오후의 일조가 차단되어 평야지의 논에서 생육한 수도보다 상시적으로 적은 일사량에서 생육하고 있는 실정이다.
이와 같이 일조량이 적은 약광조건 하에서 생육한 개체는 강광조건에서 생육한 개체에 비하여 형태적, 생리적으로 많은 차이가 있는 것으로 보고되어 있다. 형태적 반응으로서는 줄기의 도장과 비엽면적(SLA: specific leaf area)의 증가, 근량 증가의 정체, 잎 표면적의 증가, 노화 잎의 조기 고사 등을 말할 수 있으며, 생리적 반응으로는 광포화점의 저하, 잎 수명의 단기화, 근 호흡의 저하, 잎 질소함량의 증가 등을 들을 수 있다. Chonan(1967)과 Sato(1970)에 의하면 약광조건하에서 엽육조직의 두께가 얇아진다고 보고하였으며, Murata(1961)와 Sato(1970)는 차광한 수도의 잎에서 광합성속도가 저하한다고 밝힌 바 있고, 이와는 반대로 광합성속도의 증가를 인정한 보고도 있다(Kumura, 1969; Uchida et al., 1980). 보통 약광조건에서 생육한 식물의 광합성속도는 광포화점의 저하(Björkman and Holmgren, 1963; Björkman and Holmgren, 1966; Lundegårdh, 1964) 및 강광하에서의 광합성속도의 감소(Murata, 1961; Sato, 1970) 등이 잘 알려져 있으나, Kiyosui와 Tsuno(1958)는 차광처리에 의하여 엽신질소농도의 증가로 광합성속도가 향상된다고 밝힌 바 있다. 또한, 필자의 연구에 의해서도 차광처리후 대두(Lee et al., 1995a; Lee et al., 1995b)와 더덕(Lee et al., 1998a; Lee et al., 1998b)에서 광합성속도의 증가를 구명한 바 있다. 그러나, 약광조건에 대한 광합성작용의 반응은 생육시기별로 서로 다른 양상을 보인다 라는 지적도 있다(Kumura, 1969; Lee et al., 1995a).
이상과 같이 약광조건에 대한 작물의 영향이 아직까지도 부분적인 범위와 비교생리학적 연구에 국한되어 앞으로의 많은 연구와 정보의 축적이 요구되고 있는 실정이며, 우리나라 중산간지역의 수도수량생산의 안정성과 다수확재배를 위해서는 내음성품종의 선정도 매우 중요하다고 생각된다. 따라서 본 연구는 차광처리에 따른 수도의 광합성능력 변화와 약광조건에서 생육한 개체의 광 강도별 광합성속도의 변화 및 형태적 형질의 영향 등을 구명하고자 실시하였다.
II. 재료 및 방법
본 연구는 삼강벼를 이용하여 1/5,000 a 폿트에 논에서 채취한 토양을 풍건, 소독시킨 후, 폿트 상면에서 10 cm까지 충전한 뒤, 6월 13일에 균등하게 생육한 개체를 선발하여 폿트당 1주씩 이식하였다. 비료는 복합비료(N:P2O5:K2O=21:17:21)를 이용하여 pot당 10 g씩 전량기비로 시용하였으며, 폿트의 수분조절은 이식 당일부터 폿트의 상면이 항상 담수상태가 되도록 수시로 관수하였다. 기타 병충해 및 관리는 표준재배법에 따라 실시하여 비교적 양호한 생육을 보였으며, 출수기는 8월 24일이었다.
정상적으로 균일하게 생육한 폿트 10개를 8월 28일부터 9월 11일까지 15일간 인위적으로 차광처리를 실시하였는데, 차광처리방법는 하우스형 파이프를 이용하여 가로 6 m, 세로 3 m, 높이 2.5 m의 소형하우스를 제작하여 시판되고 있는 흑색 차광재료를 하우스에 덮어 그 하우스 내에 폿트를 놓아 생육시켰으며, 차광율는 56%로 조절하였다.
조사는 차광처리전 8월 28일에 광 강도별 광합성속도와 증산작용 및 기공전도도 등을 측정하였고, 엽면적, 엽록소함량, 비엽면적, 건물중 등도 조사하였다. 또한, 차광처리 종료일인 9월 11일에 무처리구와 처리구의 폿트에 대하여 4반복으로 상기항목에 대하여 조사하였다. 조사방법으로서, 광합성측정은 LA-6400 휴대용 광합성측정기를 이용하여 광합성속도, 증산작용 및 기공전도도 등을 측정하였으며, 광 강도별 광합성속도의 변화는 LA-6400의 인공광선을 이용하여 조사하였다. 또한 엽록소함량은 미놀타 SPAD 503를 이용하였고, 비엽면적은 엽면적측정기로 엽면적을 측정한 후, 80℃ 건조기에 24시간 건조시켜 건물중을 달아 엽면적/건물중의 비로 구하였다.
III. 결과 및 고찰
일반적으로 약광조건에서 생육한 개체의 잎이 얇아지는 사실은 수도(Chonan, 1967; Sato, 1970)뿐만 아니라, 콩(Lee et al., 1995a; Lee et al., 1995b), 사료작물 등 다양한 식물(Chatterson et al., 1972; Miura et al., 1983; Chujo and Asano, 1973)에서도 구명되었으며, 엽위별로도 차이가 있다는 사실도 잘 알려져 있다. 본 실험에서 차광처리에 의한 비엽면적(SLA:specific leaf area)을 조사하였던 바, Table 1에서 보는 바와 같다. 차광처리 15일 후(9월 11일)를 살펴보면, 처리구가 무처리구보다 현저하게 증가하는 경향으로 지엽에서는 61%의 증가를 보였고 그 바로 아래의 하위엽에서는 39%의 증가를 나타냈다. 그러나 지엽에서부터 네번째 잎에서는 처리간에 차이가 나타나지 않았다. SLA를 엽위별로 보면 무처리구에서는 지엽보다 그 하위엽에서 높은 경향이었고, 처리구에서는 엽위별 SLA의 차이는 뚜렷한 경향을 보이지 않았다. 이와 같은 결과에서, 일반적인 수도재배에 있어서도 상위엽보다는 하위엽에서 음엽화되기 쉬우며, 약광조건 하에서는 상위엽도 음엽화될 수 있다는 사실을 본 실험에서 알게 되었다.
표면적에 대한 차광처리의 영향을 보면 모든 잎에서 처리구가 무처리구에 비해 표면적이 증가하는 경향이었고, 엽위별로는 지엽보다 그 아래의 하위엽이 넓은 경향으로 Yoshida와 Cock (1971)의 결과와 일치하였다.
차광처리에 의한 엽록소함량의 변화를 Table 2에 나타난 바와 같다. 보통 SPAD값과 엽록소함량과는 밀접한 관계에 놓여 있으며, Y = 0.16 X + 1.3 (Y: Chrolophyll a + b, X: SPAD)라는 1차 직선회귀로 나타낼 수 있고, 고도의 정의 상관관계가 인정된다고 보고(Wada et al., 1992)된 바, 본 실험에서는 엽록소함량을 SPAD값으로 나타내었다. 차광처리 15일후의 SPAD값을 보면 무처리구에 비해 차광처리구의 모든 잎에서 높은 경향이었으며, 엽위 간에는 유의성이 인정되지 않았다. 또한, 처리전의 8월 28일과 15일 이후인 9월 11일 무처리구의 SPAD값을 비교해 보면 9월 15일의 SPAD값이 8월 28일 측정한 SPAD값보다 감소하는 경향으로 나타났는데, 이는 출수기 이후의 잎의 노화현상에 기인한 것으로 사료된다. 이상의 결과에서, 출수기 이후의 차광처리는 잎의 노화에 따른 엽록소의 파괴 및 분해를 억제하는 효과가 있는 것으로 사료되며, Wada 등(1990-1992)에 의하면 잎이 강한 광에 있으면 장해를 받을 경우가 있고, 포화광 이상의 광은 잎의 노화를 촉진시킬 가능성이 있다고 지적한 바 있으며, 출엽 후 3주 이후에 광합성 광포화점 부근의 50% 차광에서 노화가 크게 지연되었다고 보고(Wada et al., 1992)한 바, 본 실험의 결과와 동일하였다.
작물의 광합성능력은 많은 요인의 영향을 받지만, 일반적으로 포화광도 이상의 광 조건에서는 CO2 농도의 영향을 크게 받고, 수분과 무기양분의 공급정도에 따라서도 차이가 있다. 그러나, 이들 요인이 충분한 조건이더라도 포화광도 이하에서는 광 강도의 영향이 매우 크다. Gabriesen(1960)은 광 강도와 광합성능력과의 관계를 검토하였던 바, 강광조건에서는 동일한 수준의 광합성능력을 보이나, 약광조건에서는 서로 다른 광합성능력을 보이는 반면, 약광조건에서는 동일한 광합성능력을 보이더라도 강광조건에서 서로 다른 광합성능력을 보이는 경우가 있다고 보고한 바 있다. 또한, Burnside와 Böhning(1957)은 양지식물이라도 장기간 음지조건에서 생육하게 되면 음엽화 된다고 밝힌 바, 잎의 발육과정에 있어서 광 조건의 중요성을 지적한 바 있다.
본 실험에서는 출수기 이후의 차광처리에 의해 광합성속도 변화를 조사하였던 바, Fig. 1과 2에서 보는 바와 같다. Fig. 1은 차광처리전인 8월 28일에 지엽과 지엽으로부터 2번째와 4번째의 하위엽을 측정한 결과이다. 그림에서 나타낸 바와 같이 측정잎 모두에서 광 강도가 증가함에 따라 광합성속도도 증가하는 경향이었으며, PAR(광합성 유효광선) 1,000 μmolm-2s-1이상에서 거의 일정한 광합성속도를 보였고, 엽위간에는 큰 차이가 나타나지 않았다. 이보다 생육이 진행된 9월 11일에 측정한 결과를 보면 Fig. 2에 나타난 바와 같다. 이 그림에는 무처리구와 차광처리구 각각 광 강도에 따른 광합성속도의 변화를 나타내었는데, 무처리와 처리구 모두에서 광 강도가 증가함에 따라 광합성속도도 증가하는 경향으로 광 강도에 따른 광합성의 반응은 8월 28일 측정한 결과와 거의 비슷하였다. 차광처리의 영향을 살펴보면, 무처리구보다 처리구의 광합성속도가 모든 잎에서 높은 경향이었는데, 엽위별 그 증가량을 구체적으로 검토해 보면, Table 3에서 보는 바와 같다. 광합성속도는 PAR 2000 μmolm-2s-1일 때의 광합성 속도로서 지엽은 무처리구에 비해 23%의 증가를 보였고, 그 바로 아래의 하위엽에서는 59%, 4번째 잎에서는 42%의 증가를 보였으며, 8월 28일의 처리 전 수준을 유지하는 경향이었다. 그러나, 무처리구는 8월 28일에 비하여 광합성속도가 모든 잎에서 감소하는 경향이었는데, 이는 잎의 노화에 의하여 광합성능력이 감소된 것으로 사료된다.
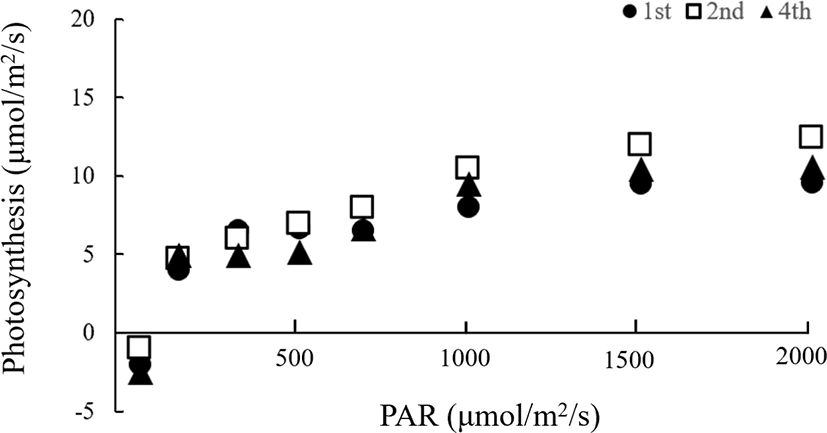
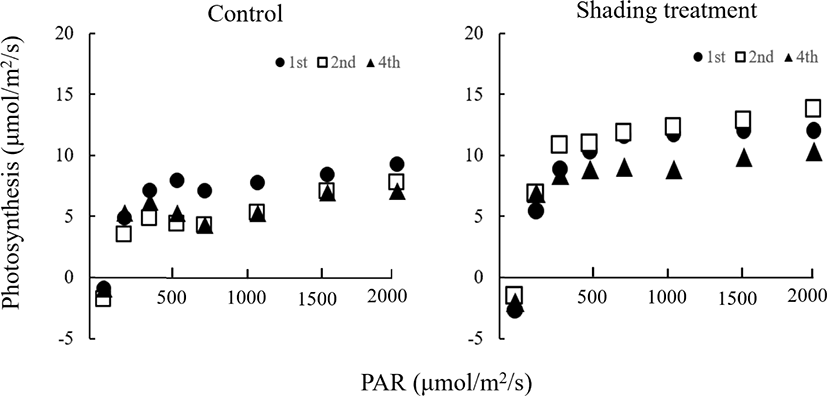
앞에서 검토해 본 바와 같이, 본 실험에서는 Murata (1961)와 Sato (1970)가 보고한 바와는 달리, Kiyosui와 Tsuno (1958), Uchida 등(1980)의 보고와 일치하였는데. 이 원인으로 Kiyosui와 Tsuno (1958)는 차광처리에 의하여 엽신질소농도의 증가로 광합성속도가 향상되는 것으로 밝히고 있다. 또한, Hidema 등(1991)은 노화과정에서의 차광은 Ribisco 함량의 감소를 적어 광합성속도가 향상되는 것으로 보고하고 바 있다.
이상의 결과를 종합 고찰하여 보면, 차광에 의한 작물의 순응은 크게 두 가지로 분류할 수 있는데, 하나는 형태적 변화에 관계없이 광환경에 대하여 광합성 관련 특성이 조절능력을 발휘하는 것이 있고, 또 다른 하나는 약광하에서 적응하여 얇고 넓은 잎을 형성하는 능력을 발휘하는 경우가 있다. 본 실험에서는 차광처리에 의하여 SLA와 표면적이 증가하는 경향으로 나타나, 잎이 얇고 넓은 경향을 보였고, 체내 성분조절로서는 엽록소함량을 증가시켜 잎의 노화를 억제하는 반응과 함께 광합성속도를 높게 유지하는 경향이었다. 이와 같이 약광조건 하에서 광합성속도가 상승하는 원인에 대해서는 이미 밝힌 엽록소함량에 의한 것으로 일시적 현상으로 사료된다.
IV. 요 약
차광처리가 수도의 형태적 형질과 광합성속도 및 그 관련형질에 미치는 영향을 조사하기 위해 1/5,000a와 그너 폿트에 1주씩 화영벼를 재배한 후, 출수기 이후에 56%로 차광처리를 15일간 실시한 후에 조사하였다. 차광처리에 의해 SLA는 모든 엽위에서 증가하였고, 하위엽보다 지엽에서 그 증가가 현저하였으며, 잎의 표면적도 처리구에서 넓은 경향이었다. 또한 엽록소 함량은 차광에 의해 모든 잎에서 처리구가 증가하는 경향이었고, 8월 28일보다 9월 11일에서 감소하였다. 광합성속도는 모든 잎에서 광 강도가 강할수록 증가하는 경향이었고, 차광처리에 의해 무처리에 비해 증가하는 경향이었다.